| 2.5 Mechanizmy na tkáňové úrovni |
|
Studium radiačních účinků na subcelulární a celulární úrovni je samozřejmě základním předpokladem pochopení účinků na vyšších úrovních organizace, jako jsou tkáně, orgány a celé tělo. Zde však je, podobně jako v jiných oblastech, zřejmé, že „celek je více než součet jeho částí". Zatímco některé tkáňové radiační účinky lze vyložit na základě toho, co je známo o buněčných mechanizmech, jiné jsou způsobeny interakcí různých tkáňových komponent a nelze je předvídat na základě toho, co bylo popsáno v předchozích kapitolách. Jedním z příkladů takových kooperativních účinků, který nyní dosáhnul širokého uznání, i když zaznamenal při prvním objevu určitý skepticizmus, je tak zvaný „bystander effect". Zatímco klasicky by bylo možno předpokládat, že radiační poškození nastává pouze v buňkách, kterými prošel foton nebo částice, studie provedené během posledních 20 let objevily necílené účinky. Jestliže jsou například jednotlivé buňky ozářeny velmi tenkým mikrosvazkem záření, vykazují sousední buňky často účinek dokonce na úrovni svých DNA. Když se k neozářeným buňkám přidá kultivační médium, ve kterém byly ozařovány buňky, odpovídají neozářené buňky stejným způsobem. Zdá se tedy, že buňky, které byly zasaženy zářením, vytvářejí difuzibilní faktory, které jsou schopny ovlivnit jiné sousední buňky - „bystandery". Je třeba zjistit, jaké jsou to faktory, avšak existuje určitý důkaz o tom, že zahrnují reaktivní kyslíkové složky. Takové účinky byly většinou pozorovány v případě hustě ionizujícího záření, například záření α.
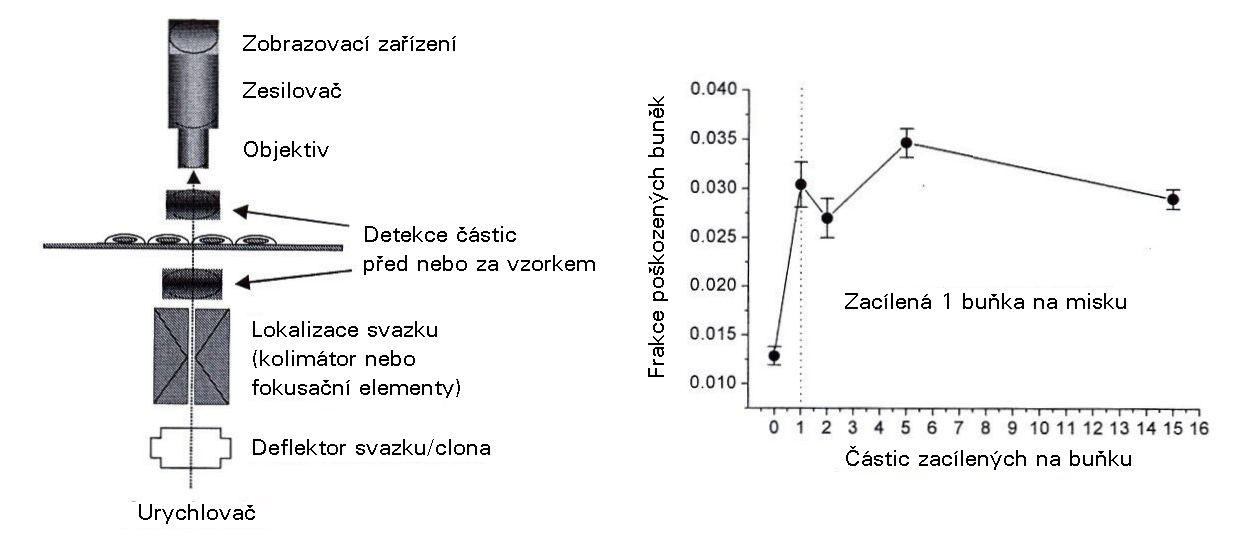
Bystander efect po ozáření mikrosvazkem. Jedna buňka na misku (obsahující několik tisíc buněk) byla ozářena, avšak frakce poškozených buněk vzrostla z 1 % (spontánně, bez ozáření) na zhruba 3% (při zaměření 1 až 15 částic na tuto ozářenou buňku) (modifikováno podle Prise et al. (2006) Dose Response 4: 263-276) Výzkumní pracovníci začali teprve nedávno pracovat s trojrozměrnými tkáňovými modely zahrnujícími různé typy buněk a strukturovanými takovým způsobem, že umožňují typ intracelulární komunikace, který by nastal in vivo, například vytvoření spojení („gap junctions") umožňujících pohyb molekul a iontů spojovacími kanály mezi cytoplazmami sousedních buněk nebo uvolňování a vychytávánírůstových faktorů a dalších cytokinů, které ovlivňují zánik buněk, proliferaci a diferenciaci. Některé z těchto signálních cest byly analyzovány dříve v monokulturách určitých buněčných typů, jako jsou fibroblasty, avšak další kroky směřují k pochopení celého spektra systémových účinků. Tuto část zakončíme zmíněním jednoho aspektu tkáňové radiační odpovědi, který do značné míry nastává účinkem na konkrétní individuální buňky, hlavně kmenové buňky, které jsou odpovědné za obnovu tkání. Tak zvaný „Zákon Bergonie a Tribondeau" (1906) uvádí, že „radiosenzitivita tkáně je přímo úměrná reprodukční aktivitě a nepřímo úměrná stupni diferenciace". Tito autoři pozorovali jako první, že nádorové buňky jsou obecně radiosenzitivnější než normální buňky těla a dále, že mezi buňkami těla jsou nejcitlivější spermatogonie a erytroblasty, epidermální kmenové buňky a gastrointestinálníkmenové buňky, zatímco nejméně citlivé jsou nervové buňky a svalové buňky. Existuje samozřejmě několik málo výjimek z tohoto „zákona", avšak vcelku lze zcela rozumně předpokládat, že buňky, které se rychle reprodukují, budou mít v průměru méně času na opravu poškození vyvolaného v jejich DNA, a proto budou více trpět následky neopravených nebo chybně opravených lézí. Proto se dokonce i určitá část tkáňových efektů vztahují zpět k tomu, co nastává během několika málo hodin po ozáření na úrovni DNA.
|