| 2.3.4 Chromozomové aberace a bodové mutace |
|
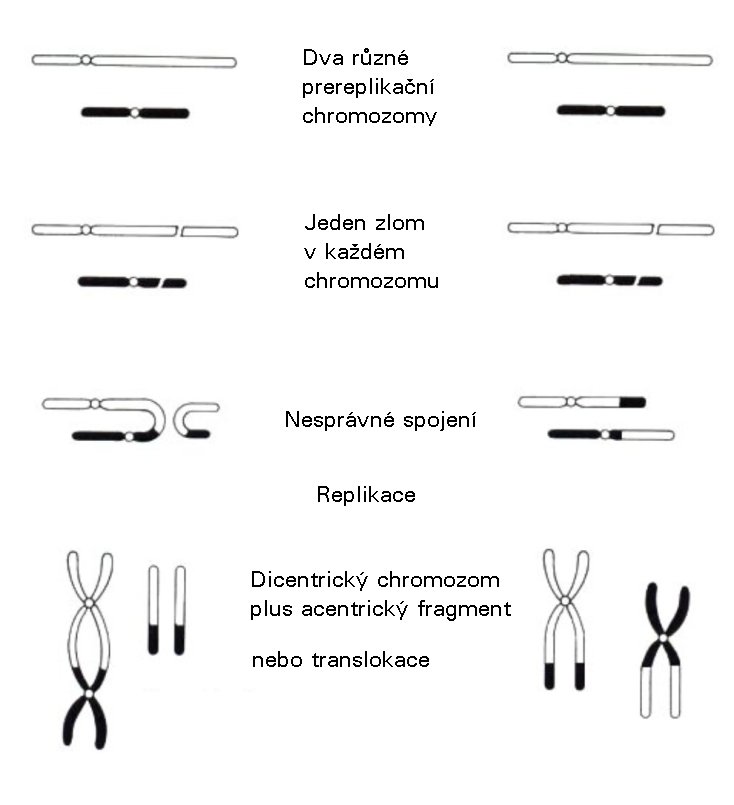
Je třeba uvažovat, že při opravě radiačního poškození se obnovuje integrita DNA a funkce buňky může normálně pokračovat. Avšak naneštěstí, i když jsou buněčné mechanizmy opravy zcela účinné, nejsou dokonalé. Existuje určitá pravděpodobnost chyb, která je samozřejmě vyšší v případech, ve kterých je třeba opravit shluky poškození. Při opravě dvojného zlomu je možná řada různých výsledků: 1) úplná oprava, to jest obnova původní sekvence DNA, hlavně úspěšným procesem HR; 2) oprava celkové chromatinové struktury s malými chybami v sekvencích DNA, jako jsou vypuštění a vložení nastávající někdy při NHEJ; 3) připojení „chybných" konců během opravy dvou současně se vyskytujících dvojných zlomů na jednom chromozomu vedoucí k větší míře delecí nebo inverzí; 4) připojení „chybných" konců během opravy současně se vyskytujících dvojných zlomů na dvou různých chromozomech vedoucí k symetrické výměně materiálu mezi těmito dvěma chromozomy zvané translokace; 5) připojení „chybných" konců vedoucí k asymetrické výměně mezi dvěma poškozenými chromozomy a tvorba tak zvaného dicentrického chromozomu (chromozom se dvěma centromerami) a dvou acentrických fragmentů.
Chromozomové aberace způsobené nesprávnou opravou dvojných zlomů různých chromozomů (modifikováno podle Hall (2000) obr. 2.6) Proces účastnící se tvorby jednoho či druhého typu chromozomálních aberací (strukturní abnormality na úrovni chromozomu) není dosud zcela vysvětlen, avšak je zde jasný důkaz existence všech těchto procesů. Poměr špatně spárovaných a úspěšně opravených dvojných zlomů závisí na mnoha faktorech, jako je typ buňky, stádium buněčného cyklu, kvalita záření a dávka. Obvykle je ten poměr řádu 1.10-5 na buňku na mGy. Protože očekáváme 4.10-2 dvojných zlomů na buňku na mGy (40 na buňku na Gy), je zřejmé, že je více než 99,9 % všech těchto zlomů úspěšně opraveno. Četnost úspěchu je dokonce ještě vyšší pro opravu jednoduchých zlomů a poškození bází. Bodové mutace (vypuštění, vložení nebo výměny jednotlivých bází) se vyskytují s podobnou frekvencí jako chromozomové aberace po ionizujícím záření, to jest zhruba 5.10-6 na buňku na mGy. Některé z nich ovšem pocházejí z chyb provedených během HR dvojných zlomů, tak že tato hodnota nám poskytuje maximální frekvenci chyb, které by bylo možno přisoudit opravě jednoduchých zlomů a modifikací bází. Při zhruba jednom z těchto dvou typů poškození na buňku na mGy (1 000 na buňku na Gy) to znamená četnost úspěchu lepší než 99,999 %. To je též konzistentní s měřeními přesnosti in vitro, se kterou polymerázy vkládají nové nukleotidy a s přesností, se kterou jsou chybně vložené nukleotidy rozpoznány a buď odstraněny ihned během procesu opravy („proof-reading") nebo opraveny později („mismatch repair"). Na tomto místě je opět vhodné připomenout statistickou povahu všech dosud popsaných procesů. Ionizace nenastává, pokud je energie přicházejícího fotonu nebo částice nedostatečná. Pokud je energie dostatečná, existuje pravděpodobnost tvorby radikálů, které s určitou pravděpodobností povedou k poškození DNA. Toto poškození nebude nikdy opraveno se 100% četností úspěchu, takže jakákoli ionizace může vést k chromozomovým aberacím nebo bodové mutaci s určitou pravděpodobností. To vše jsou důležité skutečnosti, které je třeba uvažovat při diskusi účinků malých dávek záření.
|